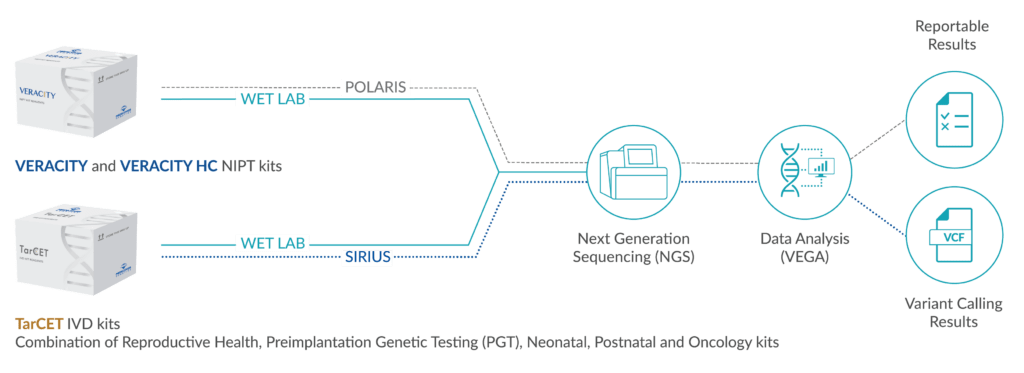
Scientific background
Intellectual disability, defined as an IQ of less than 70, has a prevalence of 1.5% to 2%; earlier data reported 2% to 3%. More severe forms with an IQ <50 have a prevalence of 0.3% to 0.4%. Males are more commonly affected due to X-linked genes. The causes o fintellectual disability are diverse; however, genetic factors are involved in at least 50% of cases. Comorbidities such as behavioral disorders and/or epilepsies are common. An accurate diagnosis is crucial for the patients and their families. Knowing the cause of the disability usually allows estimation of a prognosis, initiation of individual support measures if necessary, avoidance of further costly diagnostic measures, and statements about a possible risk of recurrence. If the cause of an intellectual disability is not clear, an empirical risk of recurrence of approximately 8% must be assumed for further pregnancies. Despite increasing numbers of new genetic syndromes associated with intellectual disability in recent years, the cause of intellectual disability still remains unexplained in some of patients.
In syndromic forms of intellectual disability, a characteristic combination of malformations, minor external abnormalities or characteristic behaviors may suggest a tentative diagnosis, which can be clarified with targeted diagnostics (e.g., fragile X syndrome, Rett syndrome, Angelman syndrome). However, many patients present with non-characteristic symptoms making the diagnosis impossible even for an experienced pediatrician or clinical geneticist. In this situation, the diagnostic approach to suspected genetic causes was global, i.e., the entire genome of the patient was investigated; however, the resolution increased over time, allowing a multitude of “new” causes to be identified. In the early days – and continuing to date – the examination starts with a chromosomal analysis, which can detect abnormal distributions of whole chromosomes, e.g., trisomies, or of smaller chromosomal parts, e.g., partial trisomies. Approximately 15% of developmental disorders are caused by chromosomal abnormalities that can be identified by light microscopy. However, even with a high resolution of 550 to 600 bands per haploid chromosome set, which can be achieved in routine diagnostics, changes that fall below 5-10 Mb cannot be detected. Therefore, a high-resolution chromosomal analysis using chromosomal microarray (CMA) is performed as a second diagnostic step. Large studies have shown that so-called copy number variations (CNVs), i.e., submicroscopic small deletions or duplications, are responsible for approximately 10 to 15% of cases of intellectual disability with inconspicuous chromosomal analysis. It has been shown that such CNVs are also frequently found in autism spectrum disorders, which occur both in isolation and in combination with a developmental disorder.
Even with these investigations, about 60% of the causes of developmental disorders remain unexplained. Since developmental disorders often occur sporadically, i.e., as an isolated case within a family, it has been assumed that new mutations occur, for example, in genes that are important for the development and interconnection of neurons, especially since humans have a high rate of new mutations.
Indeed, in recent years, several studies investigating patients with intellectual disability using new high-throughput techniques such as exome sequencing have confirmed that dominant new mutations seem to contribute to a large extent to the cause of severe (IQ < 50) intellectual disability. While the risk of chromosomal trisomies increases with maternal age, the rate of dominant new mutations increases with paternal age. Among the patients examined, variants were found in several genes. However, the causative mutation was in a gene already described in connection with developmental disorders in 16% of the patients in the study by de Ligt et al and in 35% of the patients in the study by Rauch et al. Based on these studies, it is thought that up to 50% of severe non-syndromic developmental disorders are caused by de novo point mutations and small indels with a large degree of genetic heterogeneity. Mutations in unknown genes or genes not known to be associated with developmental disorders require immense effort, including functional testing, to prove causal association. Nevertheless, exome sequencing is increasingly used in routine diagnostics, predominantly as a trio analysis, while genome sequencing is still largely limited to research. It is expected that using next generation sequencing (NGS) the underlying cause of approximately 30% of previously unexplained severe developmental disorders can be determined.
Currently, the diagnosis of a non-syndromic developmental disorder is made using a stepwise diagnostic approach starting with chromosomal analysis. If the findings are normal, a CMA is performed. If necessary, a molecular genetic examination can be conducted, e.g., to exclude fragile X syndrome or Angelman syndrome.
References
Harripaul et al. 2017, Cold Spring Harb Perspect Med 7:a026864 / Vissers et al. 2016, Nat Rev Genet 17:9 / Gillissen et al. 2014, Nature 511:344 / Musante et al. 2014, Trends in Genetics 30(1):32 / de Ligt et al. 2012, NEJM 367:1921 / Rauch et al. 2012, Lancet 380:1674 / Sharp 2011, Genet Med 13:191 / Cooper et al. 2011, Nat Genet 43:838 / Topper et al. 2011, Clin Genet 80:117 / Vissers et al. 2010, Nat Genet 42:1109 / Heinrich et al. 2009, J Lab Med 33:255 / Fan et al. 2007, Hum Mutat 28:1124 / Rauch et al. 2006, Am J Med Genet 140A:2063 / Rost and Klein 2005, J Lab Med 29:152 / Inlow and Restifo 2004, Genetics 166:235 / Battaglia and Carey 2003, Am J Med Genet 117C:3 / Leonhard et al. 2002, Ment Retard Dev Disabil Res Rev 8:117, 2002 / Curry et al. 1997, Am J Med Genet 72:46
COFFIN-LOWRY SYNDROME
Coffin-Lowry syndrome (CLS) is an X-linked dominant inherited syndrome; the incidence is estimated to be between 1:50,000 and 1:100,000. The leading symptoms in males are intellectual disability (IQ 15-60) and a characteristic appearance. The external features are a prominent forehead, wide-set eyes, downward slanting eyelid folds, broad nose with everted nasal base, full lips with everted lower lip, hyperextensible hand and finger joints, broad tapering fingers. In infancy and childhood, muscle hypotonia is also a prominent feature. The final height is usually below the third percentile. 80% of affected individuals develop progressive kyphoscoliosis, sometimes with cardiovascular complications, and many have a funnel (sunken) or pigeon chest. In addition, about 15% show mitral regurgitation, and about 30% have sensorineural hearing loss. Seizures occur in about 5%, and drop attacks, a sudden loss of tone without loss of awareness to auditory or tactile stimuli, occur in about 20%. Females are usually more mildly affected, and the spectrum of symptoms can range from mild symptoms with normal intelligence to complete penetrance as seen in the male sex, depending on X inactivation.
CLS is caused by variants in the RPS6KA3 gene on the short arm of the X chromosome (Xp22.1-22.2). The gene encodes a serine-threonine kinase. Germ cell mosaics have been described.
References
Rogers and Abidi. 2002 Jul 16 [updated 2018 Feb 1]. In: Adam MP, Ardinger HH, Pagon RA, Wallace SE, Bean LJH, Mirzaa G, Amemiya A, editors. GeneReviews® [Internet]. Seattle (WA): University of Washington, Seattle; 1993–2021 / Pereira et al. 2010, Eur J Hum Genet 18:627 / Pereira et al. 2007, Hum Genet 122:541 / Horn et al. 2001, Prenat Diagn 21:881 / Field et al. 2006, Clin Genet 70:509 / Delaunoy et al. 2006, Clin Genet 70:161 / Hunter 2002, Am J Med Genet 111:345 / Hanauer et al. 2002, J Med Genet 39:705 / Trivier et al. 1996, Nature 384:567
FRAGILE X SYNDROME
Fragile X syndrome is the most common monogenic inherited cause of intellectual disability. In contrast to other diseases with X-linked recessive inheritance, fragile X syndrome shows healthy male carriers and in some of the female patients symptoms are as severe as in males. The incidence of affected males is estimated to be 1:5164 according to a 2009 study.
The cause of fragile X syndrome is a CGG triplet repeat expansion in the non-translated 5′ region of the FMR1 gene on the long arm of the X chromosome. The most common normal alleles in the general population have a length of 29-30 CGG repeats. Alleles in the range of 45 to 54 repeats are defined as gray zone alleles (EMQN ring trials). Some instability is already present at this repeat count, regardless of the sex of the carrier; however, expansion to a full mutation in one generation has not yet been observed in this range.
Alleles with 55 to 200 CGG repeats are referred to as premutation. In females, inheritance of premutations is unstable, usually leading to an extension of more than 200 triplets (full mutation) when they are passed on to the next generation. Above this length, methylation of cytosine residues of the repeat and adjacent regulatory elements occurs, ultimately leading to inhibition of transcription and consequent failure of the FMR1 gene product. In males, the premutation is stable when passed on to the next generation. Mothers of children with a full mutation are obligate carriers with either a premutation or a full mutation. The risk of recurrence is up to 50% for affected children depending on the sex or the length of the premutation in the mother.
In the presence of the full mutation, the developmental delay is first noticeable in childhood and usually affects speech more than motor skills. The children often have slightly abnormal measurements for body length and head circumference. Occasionally, symptoms of connective tissue weakness such as hyperextensible joints and muscle hypotonia are present. Hyperactivity and autistic behaviors are characteristic features. Apart from rather large ears, other phenotype features such as an elongated face and prominent chin are not very pronounced in childhood, but characterize adults with fragile X syndrome. Postpubertal macroorchidism is often seen in males. Female carriers of the full mutation can show variable symptoms ranging from an inconspicuous phenotype (about 30%) to intellectual disability of a severity similar to that seen in males. About 20% of premutation carriers have premature menopause (FXPOI). In older premutation carriers, particularly males, more than 30% show a progressive neurological disease pattern consisting of intention tremor, gait ataxia, parkinsonism, autonomic dysfunction and dementia, which is called fragile X tremor ataxia syndrome (FXTAS).
References
Mila et al. 2018, Clin Genet 93:197 / Hall et al. 2018, Handb Clin Neurol 147:377 / Hagermann et al. 2017, Nat Rev Dis Primers 3:17065 / Pugin et al. 2017, Neurologia 32:241 / Biancalana et al. 2015, Eur J Hum Genet 23:417 / Spath et al. 2010, Am J Med Genet A 152A:387 / Leitlinien zur molekulargenetischen Diagnostik: Fragiles-X und Fragiles-X assoziiertes Tremor/Ataxie Syndrom 2009, medgen 21 / Rost et Klein 2005, J Lab Med 29:152 / Nolin et al. 2003, Am J Hum Genet 72:454 / Oostra et al. 2001, Clin Genet 60:399 / Oostra et al. 1993, J Med Genet 30:410
LUJAN-FRYNS SYNDROME
Patients with Lujan-Fryns syndrome (LFS), also known as X-linked intellectual disability (XLMR) with marfanoid habitus or X-lined syndromic intellectual developmental disorder (MRXSLF), present with marfanoid habitus, certain craniofacial features, generalized muscle hypotonia, developmental delay, behavioral problems, and nasal speech. Thus, there is clinical overlap with other connective tissue disorders such as Marfan syndrome (MFS), Loeys-Dietz syndrome (LDS), and Shprintzen-Goldberg syndrome (SGS). Inheritance is X-linked recessive, resulting in predominantly male patients, while female carriers are usually clinically inconspicuous.
Hemizygous variants in the MED12 gene, which encodes mediator complex subunit 12, are the genetic cause. Allelic disorders with MED12 variants include FG syndrome type 1 (FGS1) and X‑linked Ohdo syndrome (XLOS). Subsequently, variants in the UPF3B and ZDHHC9 genes have also been described in patients with intellectual disability and marfanoid habitus. However, these patients only partially exhibited the characteristic facial abnormalities of LFS, such as a long narrow face, prominent forehead, broad nasal root, short philtrum, micrognathia, and high palate.
References
Charzewska et al. 2018, Clin Genet 94:450 / Hackmann et al. 2016, Am J Med Genet 170A:94 / Lyons. 2008 Jun 23 [Updated 2016 Aug 11]. In: Adam MP, Ardinger HH, Pagon RA, et al., editors. GeneReviews® [Internet]. Seattle (WA): University of Washington, Seattle; 1993-2021. / Callier et al. 2013, Clin Genet 84:507 / Schwartz et al. 2007, J Med Genet 44:472 / Tarpey et al. 2007, Nat Genet 39:1127 / Raymond et al. 2007, Am J Hum Genet 80:982
RUBINSTEIN-TAYBI SYNDROME
Rubinstein-Taybi syndrome (RTS) is characterized by symptoms of low intelligence, postnatal growth delay with reduced final height and microcephaly. Distinctive facial features are seen, such as a deep hairline, broad, arched eyebrows, outwards and downward sloping eyes, base of the columella below the nostrils when seen in profile, retrognathia, and dental anomalies. Additionally, broad, often radially angled thumbs and broad big toes are seen. When laughing, there is a characteristic facial expression with almost closed eyes. Seizures occur frequently. RTS usually occurs sporadically, and the prevalence is estimated at approximately 1:100,000‑125,000.
Pathogenic variants in the CREBBP gene, which codes for the cyclic AMP-regulated enhancer binding protein, are a known cause of RTS (approximately 50-70%). In addition, pathogenic variants have been described in the EP300 gene (approximately 5%), which codes for the E1A binding protein p300. These two proteins are transcriptional co-activators involved in many signaling pathways within the cell (e.g., DNA repair, growth, differentiation and apoptosis).
References
Korzus 2017, Adv Exp Med Biol 978:39 / Fergelot et al. 2016, Am J Med Genet A 170: 3069 / Milani et al. 2015, Pediatr 41:4 / Stevens et al. 2014, Gene reviews / Bartsch et al. 2010, Am J Med Genet 152A:181 / Schorry et al. 2008, Am J Med Genet 146A:2512